:
B
Edu Cómputo A.C., UMSNH, ANUIES y SINED (2010).MX
Mayo 19, 2010
_________________________________________________
Evolución y el origen de la vida
Nuestro origen, es un desafío cognitivo extraordinario y una pregunta que siempre emociona a la humanidad, las primeras moléculas de la vida RNA y aminoácidos, son puestos a discusión en la teoría de la coevolución química; la vida apareció muy rápido en la tierra hace unos 3.8 mil millones de años, se discute si existió un antepasado común surgido por probabilidad, se reconstruye la ruta histórica más probable que siguió el surgimiento de los aminoácidos. Una teoría fundamental de la biología, sostiene que los seres vivos modernos esconden en sus genomas el árbol de la vida, que entre muchas respuestas, más en un acto de esperanza, pretenden revelar la naturaleza química, física y biológica del organismo primordial. Se cree que la lógica de los genes, permitirá a la razón perforar con audaces experimentos la incertidumbre y hacer pasos reversibles en el genoma hasta llegar a nuestro origen.
2.1. Organismo primordial
En el espacio más allá de los límites de la experiencia humana o entendimiento, algunos científicos llaman al primer ser vivo de este espacio, el incognoscible; toman este punto de vista con respecto al origen del organismo primordial. El origen del organismo primordial es por consiguiente el equivalente biológico de la teoría del “Bing Bang” en astrofísica; en la que astrofísicos piensan que el universo entero era una vez del tamaño de una pelota de golf que entonces explotó para crear el universo observado. Ellos pueden rastrear los fenómenos cósmicos observados en otras galaxias, el corrimiento al rojo, y la radiación de fondo, pero ellos admiten que es “el incognoscible” acerca de cómo la pelota de golf llegó allí.
Las cianobacterias han tenido un papel fundamental en la historia de la vida en la Tierra, siendo los primeros organismos en llevar a cabo la fotosíntesis oxigénica, que cambió la química de la atmósfera y permitió la evolución de Eukarya aeróbica, sin embargo, estamos muy lejos del organismo primordial, el origen de la vida no es algo trivial como ya lo discutimos en el apartado anterior ¿Qué es la vida? Todos hemos tenido curiosidad por nuestro origen mediato y el primitivo, sin duda los recientes adelantos científicos no prometen determinar cómo fue el organismo primordial
[i]
,
[ii]
,
[iii]
,
[iv]
y cómo llego a nuestro universo o se desprendió de él. Sin embargo este trabajo intenta aportar toda una serie de enigmas que inquietan al espíritu humano, bajo la tesis que los organismos fósiles y modernos, esconden la arquitectura primordial.
Por ejemplo, la mayoría de las bacterias pueden haber evolucionado en las tierras de un antepasado común, de acuerdo con un análisis realizado por Matt Kaplan, donde las relaciones entre las diferentes familias de bacterias ha sido polémica; tradicionalmente, los árboles filogenéticos que muestran cómo las bacterias están relacionadas unas con otras se han basado en dos técnicas diferentes, arrojando resultados diferentes cada una. Un árbol se construye mediante la comparación de los genes que codifican ARN ribosomal, mientras que el otro método utiliza entre 20 y 40 genes esenciales que se encuentran en casi todos los organismos vivos.
[v]
El camino de la evolución de las bacterias fotosintéticas anoxigénicas a las cianobacterias oxigénicas es discontinuo en cuanto a la fotoquímica (sistemas de reacción fotofísicas). Es difícil describir este proceso de transición, simplemente porque no se reconocen los organismos intermedios entre los dos grupos de bacterias. El Violaceus Gloeobacter PCC 7421
[vi]
generalmente es un organismo modelo adecuado para el análisis, ya que aún posee características primordiales tales como la ausencia de las membranas tilacoides. Todo el análisis del genoma, de bioquímica y de inspección biofísica del violaceus ha favorecido la hipótesis de que existió un organismo intermedio. Por otra parte, la segunda técnica nos dice que la diferenciación de especies es un proceso evolutivo que podría ser impulsado por los cambios en un pequeño número de genes, y este proceso podría dar información de los detalles del control de diseño.
El campo de la biología molecular ha producido una visión adicional a las preguntas del origen
[vii]
, por ejemplo: ¿es posible que el origen de la vida se dio en la “Tierra” por coevolución química? Nosotros los ingenieros de la computación y la electrónica, hemos mostrado interés en la genética debido a las similitudes entre los procesos genéticos e informáticos. Hay un código genético digital y hay lógica digital con funciones y matrices lógicas. Hay incluso “errores que corregir”
[viii]
en el copiado digital o en la codificación genética.
[ix]
,
[x]
La pregunta del cómo surge tal código genético no es menor a la interrogación del cuál es la conexión racional de este código con el organismo primordial.
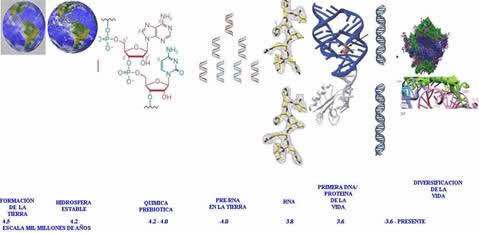
Fig. 1. Un panorama a la evolución de la vida.
El intento de hacer predicciones acerca de la vida en otros lugares sobre la base de observaciones sobre la Tierra, es inherentemente difícil debido al tamaño de muestra 1. También se llena de polémicos conceptos "antrópicos"
[xi]
. Sin embargo, no hay razón en principio del por qué no se puede hacer esto. Si usamos la teoría de probabilidades de incertidumbre del modelo Jaynes
[xii]
y datos sobre la vida en la Tierra sin información acerca de la vida extraterrestre, y a continuación, la teoría de la probabilidad nos devolverá un campo de distribuciones, lo que nos indica una gran incertidumbre.
Es un hecho sorprendente de que la vida surgió en la Tierra muy rápidamente después de su formación, aproximadamente hace 3.8 mil millones de años
[xiii]
y al final de una probable fase de esterilización debido a los impactos frecuentes de meteoritos; se ha utilizado este argumento para la abiogénesis (véase fig. 1). Lineweaver & Davis (crean el modelo L&D)12 han modelado este razonamiento con la teoría de probabilidades y concluyeron con una confianza del 95% (probabilidad Bayesian) de que la probabilidad de la abiogénesis en un planeta como la Tierra es mayor del 13%. Un 13% se hizo mediante un modelo donde no había peligro constante (posibilidad de que surgiera la vida por intervalo de tiempo discreto) q. La distribución de probabilidad para el tiempo tL(tL corresponde a
 ) en la que la vida surge depende de q, y esto se calcula condiciendo a tL inferior a la edad de la Tierra. Esta distribución de probabilidad es una función de verosimilitud para q cuando el tL real observado se sustituye en la misma. Combinado con una distribución previa para q, entonces podemos hacer inferencias acerca de su valor.
) en la que la vida surge depende de q, y esto se calcula condiciendo a tL inferior a la edad de la Tierra. Esta distribución de probabilidad es una función de verosimilitud para q cuando el tL real observado se sustituye en la misma. Combinado con una distribución previa para q, entonces podemos hacer inferencias acerca de su valor.
Si bien es posible e interesante para el cálculo de tales cosas, el modelo L&D contiene un error que hace que la conclusión sea válida. Lamentablemente, la conclusión antes citada depende de una elección de la distribución de probabilidad a priori sobre q, es un exceso de confianza y poco realista sobre nuestro estado de conocimiento sobre la abiogénesis. Sin embargo, este modelo parte de la ignorancia inicial, muy propia de los modelos Bayesianos.
Supongamos la existencia de un planeta que es idéntico a la Tierra primitiva (cuando las condiciones son aptas para la vida; llamamos a este tiempo to) en función de todos los parámetros microscópicos: masa, temperatura, composición química, distancia del Sol (con un sol idéntico) etc., por supuesto que este modelo sólo se aplica a los planetas similares al nuestro. Si bien esto puede parecer restrictivo, en base a nuestro conocimiento de lo real observado en la Tierra, resultan no relevantes los planetas distintos al nuestro. Imagine que se nos da el valor de una constante, µ, que es el tiempo de espera para expectativa de la abiogénesis con las condiciones anteriores. A partir del análisis de supervivencia estándar, 1/ µ es proporcional a la probabilidad por unidad de tiempo que el evento suceda, y desempeña el mismo papel de q en L&D. Es entonces que informa para nuestra sorpresa cuales fueron los eventos que ocurrieron en el planeta Tierra para el organismo primordial:
Proposición S: en el tiempo t= to (el tiempo presente. A partir de entonces un pájaro carpintero de .1 kg existe cada vez que un valor específico es requerido) existe una persona llamada Felipe Calderón y es el presidente de México en el planeta Tierra.
La vida surgió por primera vez en el planeta en un momento en tL: obviamente tL< to.
Mientras que la proposición S puede parecer demasiado específica, es la más probable para hacer inferencias correctas por el condicionamiento de un argumento que es más específico, por ejemplo, "la vida inteligente se plantea", como efecto antrópico.
Nuestras predicciones se darán en forma de distribuciones de probabilidad para todos estos parámetros. Las distribuciones de probabilidad son elegidas para representar a nuestro estado incierto del conocimiento (el marco bayesiano Jaynes). Las probabilidades de proposiciones las denotaremos por una P() y funciones de densidad de probabilidad (PDF) para las variables de un caso p().
En una distribución de muestreo: si sólo sabemos que la abiogénesis tiene una escala de tiempo µ, nuestra predicción para tL sería descrita por una distribución exponencial:
(1)
Tenga en cuenta que esto no es una suposición acerca de cualquier distribución de frecuencias que se producirían en una población de la Tierra, es sólo la distribución de probabilidad más conservadora que tiene el valor esperado μ
[xiv]
. Cuando nos enteramos de que S es cierta para el planeta que estamos viendo, la distribución revisada se trunca entre t = 0 y t = t0:
(2) y (3)
Técnicamente, esto debería haber sido calculado a partir del teorema de Bayes:
(4) y (5)
Donde el primer término en el numerador de la ecuación 1 vendría a ser el fácil. El otro término sería muy difícil de cuantificar, sin embargo, cualquier efecto además de los evidentes efectos de truncamiento (S no puede ser cierto a menos tL <t0) complicaría más el cálculo.
Por ejemplo, el hecho de que es muy poco probable que S sea verdad si tL está cerca de t0 que corresponde a la "no-observabilidad de la abiogénesis reciente" y seguiría el modelo con el factor de p (S | tL, μ). Otro posible efecto es que hay varias épocas de la historia para cualquier planeta similar a la Tierra, y las condiciones son adecuadas para que la vida se origine solamente en una de esas épocas. Sin embargo, el truncamiento simple de la ecuación 2 es suficiente para repetir la mayor parte del argumento L&D, haciendo hincapié en nuestro punto de desacuerdo con él.
El hecho de que la vida surge sorprendentemente poco después de la formación de la Tierra puede ser usado como evidencia para la hipótesis de que la abiogénesis debió ser fácil, (pensando que la vida surge en la tierra) y por lo tanto, apoyaría la conclusión de que la vida es común en el universo. Sin embargo, la evidencia no es tan concluyente como se ha dicho cuando aumentamos los factores en la ecuación. La vida es extraordinariamente rara en el universo, tal vez sólo en la Tierra, y observamos abiogénesis antes de tiempo debido a la casualidad (que tendría que ser moderadamente suerte). De ahí, a menos que haya una detección directa, la respuesta a la pregunta de siempre de "estamos solos", sigue siendo, "nadie lo sabe".
La evolución, es el concepto que pretende explicar que toda la vida en el planeta se deriva de un antepasado común. Es la hipótesis que pretende ligar todos los fenómenos físicos, químicos, biológicos por transformaciones sucesivas más o menos graduales y continuas de una sola realidad primaria.
[xv]
La bioquímica de organismos vivientes es una colección de estrategias exitosas acumuladas a través de billones de años de experimentación de la vida. La absorción de los aminoácidos en los minerales y su condensación en las condiciones que se asemejan a los de la tierra prebiótica es un tema que introdujo el famoso experimento prebiótico de Stanley Miller en 1953
[xvi]
. Sin embargo, los aminoácidos que se deben utilizar en estos experimentos es todavía una cuestión abierta. Nos preguntamos si había dos fuentes de aminoácidos para la Tierra prebiótica: (1) exógenos - el sentido de que los aminoácidos fueron sintetizados fuera de la tierra y se entregan a nuestro planeta por las partículas de polvo interplanetario, meteoritos, cometas, etc y (2) endógeno - es decir, que se sintetizaron en la tierra en las mezclas atmosféricas, fuentes hidrotermales, etc. En los estudios de la química prebiótica, el uso de una mezcla de aminoácidos, tanto endógenos y exógenos, se sugiere que el aporte exógeno de aminoácidos a esta mezcla es muy diferente de la composición media de las proteínas, y contiene varios que no son aminoácidos de proteínas. Por otra parte, la mezcla de aminoácidos a partir de fuentes endógenas se parece asemejar más a la composición de aminoácidos de las proteínas terrestres.
[xvii]
Es esencial para el origen espontáneo de la vida en la tierra, la disponibilidad de moléculas orgánicas como bloques de construcción. La famosa "sopa prebiótica" del experimento de Stanley Miller había demostrado que los aminoácidos, los bloques de construcción de las proteínas, surgieron de entre otras moléculas orgánicas pequeñas de forma espontánea en el laboratorio, por el que provocó una mezcla de metano, hidrógeno, amoniaco y agua. Estas condiciones se supone que simulan las de la tierra primitiva. Ya en 1922 Oparin propuso que la Tierra primitiva tenía una atmósfera reductora (en su clásico "El Origen de la Vida" de 1936, amplió estas ideas). Las observaciones de Júpiter y Saturno habían demostrado que contenía amoníaco y metano, y grandes cantidades de hidrógeno se infieren a estar presentes también allí (ahora se sabe que el hidrógeno es el principal componente de la atmósfera de estos planetas). Estos ambientes de los planetas gigantes eran considerados como remanentes de la captura nebulosa solar y la atmósfera de la Tierra primitiva fue asumida por analogía de manera similar. Sólo en una atmósfera reductora como esta, la síntesis de moléculas orgánicas - también los azúcares y bases orgánicas, bloques de construcción de nucleótidos - habría sido posible en gran cantidad
[xviii]
. Más tarde la investigación había arrojado dudas sobre la existencia de una atmósfera reductora, y sugirió un ambiente neutral en su lugar
[xix]
. El problema de este modelo teórico de la existencia de una sopa prebiótica es sin duda la falta de cimientos sólidos para determinar cuál era la composición química exacta de la tierra primitiva.
Un camino alternativo es la evidencia fósil. La mayor estimación de los años que la vida a estado en la tierra, se calcula en 3.85 mil millones de años.
[xx]
Esto es basado en las proporciones de isótopo de carbono en algunas de las piedras sedimentarias más viejas conocidas en la tierra, se trata de las rocas Itsaq, la piedra génesis localizada en el sur oriental de Groenlandia. Estas piedras no contienen microfósiles visibles, pero las células vivientes preferencialmente incorporan el isótopo más ligero del carbono C12 no el C13 o C14. Material que se ha originado de los seres vivientes, tiene una proporción de estos isótopos de carbono que refleja el decaimiento de los isótopos más pesados. El carbono de materiales no orgánico-biológico tiene una proporción diferente. Las proporciones de isótopo de carbono vistas en estas piedras antiguas de 3.85 mil millones de años parece como si tuvieran un origen en células vivientes. Un decaimiento similar de isótopos de carbono se ha reportado para un meteorito marciano que se pensó contenía microfósiles
[xxi]
.
¿Qué significa eso para el origen de la vida? El planeta se formó hace aproximadamente 4.5 mil millones de años y se piensa que la superficie se encontraba fundida o bajo el bombardeo continuo del espacio antiguo hasta hace aproximadamente 4 mil millones de años. Los impactos de meteoros y la actividad volcánica habrían hecho la superficie incapaz para la vida. La existencia probable de vida promedia 3.85 mil millones de años, cuando la vida casi comenzó en el planeta. Por consiguiente, el origen de vida en la tierra fue muy rápido. Los microfósiles más viejos evidencian células que se parecen a la cianobacteria que viene en una chert -una piedra parecida a la pedernal, consistiendo esencialmente en una gran cantidad de chalcedony fibroso con cantidades más pequeñas de cuarzo criptocristalino y la sílice amorfo- de Ápice de arcaica, siendo el eón más primitivo de la historia geológica o el sistema correspondiente de piedras, -eón: unidad de tiempo geológico, equivalente a 1000 millones de años- de Australia occidental fechada aproximadamente 3.43 mil millones de años.
[xxii]
Como ejemplo tenemos los estromatolitos, que son fósiles que muestran los procesos de la vida de las cianobacterias (anteriormente llamadas algas azul-verdes)
[xxiii]
.
Para el investigador mexicano Beraldi, los estromatolitos son estructuras órgano-sedimentarias laminadas (principalmente de CaCO3) adheridas al sustrato, producto de la actividad metabólica de microorganismos (principalmente cianobacterias o algas cianoprocariotes), aunque también las clorofitas participan en la precipitación de carbonatos. Son estructuras rocosas y porosas, de superficie rugosa-gelatinosa, producto de las secreciones mucilaginosas. Existen estromatolitos en cualquier era geológica (desde el Precámbrico), incluso actualmente siguen creciendo en muchos lugares del mundo. En México pueden encontrarse en la actualidad estromatolitos en la laguna de Alchichica. Puebla, en Las Huertas; Morelos, en Cuatrociénegas, Coah.; Lago de Pátzcuaro zona este, Michoacán y en otras localidades de Oaxaca, Yucatán y San Luis Potosí.
[xxiv]
Nosotros asumiremos esta evidencia fósil, que fecha a la vida en la tierra hace 3.85 millones de años. Los microfósiles estromatolitos, que muestran formas similares a las cianobacterias modernas, sugieren que la vida evolucionó en una forma similar a la bacteria de hoy y que las bacterias han cambiado poco por lo menos en los últimos 3.5 mil millones de años. Además, todos presentan la vida hoy en día, basada en una molécula de información ácida nucleica ARN (o RNA por sus siglas en inglés) que contiene información en código, necesaria para hacer una célula viviente
[xxv]
,
[xxvi]
. La información es codificada en un código polipéptido y aunque hay algunos ejemplos de variaciones ligeras en este código, ningún cambio radical existe en él. Esto es "universal", el código se interpreta por proteínas, a través de una maquinaria compleja llamada ribosomas que también son compartidas en común entre todas las cosas vivientes. Se conservan hoy estos rasgos principales de almacenamiento de información y recuperación, proporcionando la evidencia convincente que toda la vida en la tierra tiene un origen o porciones de RNA en un antepasado común.
Desde que la vida empezó, ha estado cambiando en las direcciones permisibles. Los constreñimientos físicos en la química de la vida incluso las propiedades del agua, la naturaleza del carbono y otros aspectos importantes de la biología, han permitido las variaciones en el tema original, pero sólo dentro de ciertos límites. Sin embargo, 3.85 mil millones de años son un tiempo largo y muchas variaciones han estado siendo probadas varias han tenido éxito en la naturaleza. Por ejemplo, la molécula mensajera RNA tiene que transmitir la información a través del tiempo por el cianotipo de una célula
[xxvii]
, esta es la información que ha cambiado en el tiempo. Esta molécula se ha copiado billones de veces, pero no sin algunos errores que se arrastran hasta hoy, nosotros podemos hacer aparecer los tiempos pasados, comparando las secuencias de los nucleótidos y las secuencias traducidas de las proteínas de los organismos contemporáneos. Por la cotización de las diferencias entre las secuencias y haciendo algunas asunciones modestas sobre las proporciones de los cambios en las secuencias, nosotros podemos estimar, cuándo los organismos diferentes divergieron entre sí. Los organismos muy similares, tienen secuencias muy similares, y los parientes más distantes tienen más diferencias en su código. Éste es el concepto del reloj molecular. Con esto, surge en la mente de los científicos la idea de poder usar las secuencias para construir el árbol de la vida con ramas que representan las diferentes especies. Las relaciones entre los organismos pueden ser las marcas de la evolución. Si bastantes organismos son incluidos y la mayoría de sus secuencias fueran usadas, podría construirse un árbol de la vida. Ésta es una genealogía de organismos del presente, y es muy interesante observar que ocurrirá con los proyectos sobre el genoma en este sentido, quizás nos esperan grandes sorpresas, uno de las bancos de datos que encierra estas respuestas ya se construye, “dbSNP“.
[xxviii]
En este semejante árbol, las ramas siempre divergen, ellas no unen atrás, porque las especies no se unen, excepto en eventos muy raros. El lugar dónde dos ramas vienen juntas es un tiempo, un punto cuando ellas eran las mismas especies. Más lejos y más lejano atrás en el árbol, las divergencias son más profundas y más antiguas. Si nosotros regresamos, bastante lejos estas ramas más distantes encontraríamos al antepasado común. Un sólo organismo celular que dio lugar a toda la vida en el planeta, nuestro “padre biológico primigenio”.
Construir un árbol así no es trivial. Se debe tener algún cuidado para escoger las secuencias correctas, porque no todas las secuencias son apropiadas para este trabajo. Incluso en un sólo gen, no todo el gen es útil para esta tarea. Frecuentemente, sólo la mayoría de las partes de un gen se ha incluido en la construcción del árbol de la vida. Con suerte, los rasgos más antiguos comunes a toda la vida son los candidatos a comparar. Esto se ha hecho más a menudo con el ARN ribosómico, debido a que desde que la vida tiene esta molécula se piensa que se replicó con efectividad. Ésta es la base del proyecto del banco de datos de ARNr que se lleva dentro del proyecto del Genoma Humano
[xxix]
.
Está claro que este árbol tendría tres divisiones principales. Éstos se han llamado dominios. En una jerarquía de vida, los dominios son más altos que los reinos. Los tres dominios son: bacterias, arqueobacterias y eubacterias.
Las bacterias y arqueobacterias son ambas procariotes, sin un núcleo. Ellas son diferentes, ya que aunque ellas son procariotes, es incorrecto clasificarlas juntas. De hecho, la mayoría de la comunidad científica acepta la versión de este árbol, que muestra la archaebacteria al ser relacionada más estrechamente a la eubacteria, pero éste es un problema aún debatido.
La primera célula existió ciertamente en el antepasado común. Nosotros no sabemos cuánto tiempo pasó antes de que este rasgo se hendiera en la vida. Nosotros podemos comparar los tres dominios y podemos hacer algunas suposiciones sobre lo que el antepasado común fue. Rasgos que están presentes en todos los tres dominios estaban probablemente presentes en el antepasado común. Es difícil ir más allá de ese punto, excepto en condiciones muy particulares. Para comprender mejor el concepto del árbol de la vida, hagamos un corto viaje por la lógica de la genética.
Todos los organismos vivientes generalmente tienen un código genético representado por la secuencia de nucleótidos en su ADN. Hay cuatro posibles bases subsecuentes usadas en la construcción del código denotadas por: A, G, C, y T, cada letra en el código lleva dos partes de información. El ADN normalmente está en la forma geométrica de doble hélice, donde la segunda hebra es complementaria a la primera hebra. Es decir, en la segunda hebra es una secuencia como “AGCTTT”, se reemplaza por “TCGAAA” qué lleva la misma información. Los pares de bases A-T y C-G constituyen los escalones de la espiral de ADN o ácido desoxirribonucleico, elemento básico de todo ser vivo conocido. Al leer la doble hélice, se puede lograr interpretar el código de la vida y sus secuencias de enfermedad. De ser posible estirar el ADN de una célula humana, éste mediría en promedio dos metros. Sólo el 3% del total del genoma humano está compuesto por genes - el resto son secuencia de redundancia o no funcionales a veces llamadas "desechos”. Los genes son secuencias especiales de cientos o miles de pares de bases que constituyen la matriz para la fabricación de todas las proteínas que el cuerpo humano necesita y determinan las características hereditarias de la célula u organismo. El número total de genes que existe en cada célula humana no se conoce con precisión, aunque se han identificado entre 25 y 27,000.
[xxx]
,
[xxxi]
Todos ellos, conjuntamente con el restante material genético que aparenta ser no funcional, se distribuyen en "cápsulas" llamadas cromosomas. Cada ser humano cuenta con 23 pares de cromosomas, proviniendo un juego del padre y otro de la madre. El total de 46 cromosomas humanos se encuentran en el núcleo de cada célula del cuerpo humano (excepto las células reproductoras, que sólo tienen la mitad). De esta forma, la mayoría de las células contienen toda la secuencia del modelo para crear un ser humano (ver http://genome.wustl.edu/). Cada una de las células de nuestro cuerpo se "especializa" en realizar determinada tarea de acuerdo con las instrucciones genéticas incluidas en el genoma. El resultado: la formación de sangre, músculos, huesos, órganos,... El cuerpo humano está integrado por un total de 100 billones (millones de millones) de células. Las tareas de secuenciación han sido divididas para las 3 200 Mb, en proyectos por cromosomas, citando para el cromosoma 1 un total 263 Mb
[xxxii]
y aproximadamente 35Mb en los cromosomas 21q y 22q.
[xxxiii]
Así la terminología “par base” se refiere a una letra del código genético representada por la base y su complemento, equivalente a dos bits de información en el lenguaje de la computadora
[xxxiv]
. Con esta lógica, los genetistas, Craig Venter crea un organismo unicelular, parcialmente estructurado por el hombre, con una cantidad mínima de genes que son necesarios para la vida
[xxxv]
. Por ejemplo, en el cromosoma 21, se identificaron causas genéticas de los más frecuentes casos de retraso mental significante que afecta a 1 en 700 nacimientos vivos. Se tienen 33,546,361 pares de bases (Mbp) de ADN con una exactitud muy alta. Los rasgos estructurales identificados incluyen duplicaciones que están probablemente envueltas en las anormalidades cromosomáticas y replican las estructuras en los telómeros y regiones del pericentromero. El análisis del cromosoma reveló 127 genes conocidos, 98 genes más y 59 seudo genes
[xxxvi]
.
Los humanos tienen un código genético -“el genoma”- de aproximadamente 3.3 mil millones pares de bases (6.6 gigabits o 825 megabytes)
[xxxvii]
. Los genomas humanos encajarían fácilmente en una unidad de disco duro portátil típica. Cada humano tiene dos copias del genoma, virtualmente cada célula tiene la información sobre la herencia que puso cada uno de los dos padres. Realmente los varones tienen 2% aproximadamente menos código en uno de sus genomas porque ellos sólo tienen un cromosoma “X”. Durante el crecimiento y la vida normal de un humano se procesan células, leídas una y otra vez, e interpretando los códigos genéticos, copias de varias partes pequeñas del código, y usa las copias como plantillas en la fabricación de proteínas.
Los primeros naturalistas pensaron que los rasgos genéticos se heredaban más o menos algo “análogo” a la moda estadística en que la descendencia tenía un promedio de las características de sus padres. Gregor Mendel fue el primero en comprender a través de extensos experimentos al engendrar guisantes que en el nivel más bajo de la herencia es binaria, y que hay una unidad mínima de herencia ahora conocida como “gen”. Mendel encontró que algunos rasgos son “recesivos” -dícese de los caracteres hereditarios que no se manifiestan en el fenotipo del individuo que los posee, pero que pueden aparecer en la descendencia de este-. Él también encontró que esa herencia en un rasgo, es independiente de herencia de otros rasgos
[xxxviii]
.
Se sabe que los genes llevan a cabo las secuencias de código genético para que las células específicas produzcan una proteína particular en un momento particular. Es un número esencialmente infinito de posibles moléculas de proteínas diferentes que depende del orden particular de las moléculas del aminoácido. El código para la producción de la proteína ha sido “roto” para que nosotros sepamos ahora que una sucesión de tres-letras -un codon-
[xxxix]
se usa para especificar un aminoácido particular (hay 20 aminoácidos esenciales). Por ejemplo, la sucesión que GGC, específica que la glicina del aminoácido será agregada a una molécula de la proteína. La salida detiene los codons que marcan el principio y el extremo de una proteína que codifica la sucesión, de una manera sorprendente se asemejan a los esquemas de comunicaciones digitales de datos modernos del modelo de capas ISO para las comunicaciones de computadoras en red. Hay 64 posibles codons y sólo 20 posibles aminoácidos para un poco de redundancia por si el error existe. Las secuencias del código regulador en genes que especifican las partes del cuerpo y/o a que tiempo una proteína se producirá, es mucho más complejo de ser entendido.
El tamaño del cromosoma y su número pueden variar ampliamente entre los organismos estrechamente relacionados. Esto propone un desafío para los genetistas evolutivos que están intentando dar sentido a la estructura del genoma. Un tercio de genes humanos se relaciona claramente a aquellos encontrados en las plantas, siendo en los mamíferos por lo menos 90% genéticamente similar a nosotros. Las ovejas tienen 27 pares de cromosomas; el ciervo de la India muntjac tiene simplemente 3. Nosotros tenemos unos 3.3 mil millones de pares de bases ADN; una amiba llega a tener más de 600 mil millones.
Aunque todavía es temprano, las comparaciones de secuencias de especies diferentes, sugieren que los eventos son como los estallidos de actividad de “genes saltadores”, duplicaciones genéticas y fusiones del cromosoma que juega un papel importante en la evolución. Lejos de ser una masa de basura ADN que sostiene una carga pequeña pero preciosa de genes, los genetistas están empezando a ver los cromosomas como fases muy dinámicas en que los procesos evolutivos importantes están presentes. Por ejemplo, los elementos móviles conocidos como transposons están entre las fuerzas más poderosas que forman la evolución del cromosoma. Éstos ‘genes saltadores’ llevan las instrucciones para su propia resección, duplicación e inserción en el genoma. Parece que, en ciertos periodos en la historia evolutiva, la actividad del transposon se hizo extensible como los acordeones en los cromosomas. Como resultado, la mayoría de los cromosomas hasta ahora contienen los remanentes silenciosos de los transposons.
Los genetistas evolutivos pueden hacer ejercicio de cuánto tiempo hace de un transposon, se inmoviliza mirando la acumulación de mutaciones en sus secuencias, flanqueando características. Tales estudios han sugerido que una agitación de la actividad del transposon doblo el tamaño del genoma de maíz de 1.2 mil millones a 2.4 mil millones bases hace 3 millones de años
[xl]
. En la evolución humana, los transposons, elementos esparcidos en el tiempo –Líneas-, se ha extendido a aproximadamente 100,000 copias en varios estallidos encima de los últimos 100 millones de años, el más reciente evento que ha ocurrido fue hace 25 millones de años en la antigüedad
[xli]
. Las líneas contenidas ahora son el 15% del ADN humano.
¿Por qué los elementos móviles deben dispersarse en los estallidos? Una idea intrigante, es que la mayoría de las células en el tiempo reprimen la actividad del transposon -una estrategia sensata, dado que un gen puede desactivarse si un transposon se mete en su secuencia-. Pero los costos y beneficios pueden cambiar durante los periodos de tensión evolutiva. Las proporciones aumentadas de transposición podrían seleccionarse entonces para ayudar a los organismos a adaptarse en tiempos pendencieros aumentando la variabilidad genética.
Los transposons no son el único tipo de ADN que puede reproducirse. Si hay una cosa en que el ADN es bueno es copiándose. Y como resultado, duplicaciones que van de los centenares de bases al complemento entero de la célula en sus cromosomas han configurado en la evolución los genomas modernos.
Las duplicaciones más simples producen secuencias repetidas adyacentes que son todas orientadas de la misma manera. Las longitudes de este tándem, en sus repetidas pueden variar enormemente, y a menudo se reproducen los genes enteros. La copia extra puede aumentar las mutaciones, entonces a menudo esto es inútil, pero una nueva y útil función también puede surgir. Se considera ahora que la duplicación del gen es el más probable origen de racimos de genes en que la selección natural ha formado copias de un gen original para asumir funciones diferentes. Nosotros debemos nuestro sentido del olfato y nuestro alcance de olor, por ejemplo, a la duplicación y diversificación de genes del olfativo receptor.
[xlii]
La necesidad de reproducir ADN para conocer su plantilla origen hace que la tecnología informática se desarrolle para dar las herramientas necesarias para la bioingeniería. El genoma humano contiene los pedazos cortos y gruesos reproducidos de ADN, cientos de kilobases en los extremos opuestos de un cromosoma o en el total de los cromosomas diferentes.
[xliii]
Evan Eichler, investigador del genoma de Case Western Reserve University in Cleveland, Ohio estima que por lo menos 5% del genoma humano surgió a través de esta clase de duplicaciones
[xliv]
.
Para recordar lo que ya dijimos sobre la compleja lógica genética, recordemos que, los humanos contenemos aproximadamente 27,000 genes en 23 cuerdas separadas de ADN conocidas como los cromosomas (46 si se cuentan ambos juegos del código). El número de cromosomas no es indicativo de complejidad. Los perros tienen 78; los caballos tienen 64; los helechos tienen 512.
[xlv]
El Proyecto Internacional del Genoma Humano (HGP) ha completado una preliminar secuencia del genoma humano entero del código genético. Se forman las secuencias de un número pequeño de otros organismos como el ratón, mosca de fruta, y coli. Teniendo la secuencia es muy diferente a entender lo que significa ésta.
En el futuro, los científicos notaran sin duda las desviaciones sutiles del modelo de herencia predicho por Mendel. Específicamente, la herencia de ciertos rasgos no era completamente independiente de otros rasgos. Nosotros sabemos ahora que la herencia de rasgos sólo será independiente si ellos se llevan en cromosomas diferentes y que la probabilidad de heredar los rasgos juntamente llevados por el mismo cromosoma es proporcional a la distancia física entre los dos genes en el cromosoma. Los estudios de herencia han producido mapas del genoma que muestra la situación aproximada de algunos genes de rasgos y genes de enfermedades genéticas humanas en los cromosomas específicos. Esta información puede combinarse en el futuro con los datos de la secuencia detallados para descubrir los genes que son responsables para las enfermedades genéticas. Hay una estimación de 3000 enfermedades genéticas humanas diferentes.
El código genético se ha comparado a un cianotipo que especifica el plano de un organismo. De hecho el código genético no sólo específica el plano del organismo, además mantiene los mecanismos necesarios para “leer” el código y fabricar los componentes del organismo, así como específica los procedimientos necesitados para los procesos de vida del organismo acabado. Los organismos simples son genéticamente definidos completamente. Cada gusano del nematodo diminuto tiene 958 células exactamente. Los humanos, por otro lado, tienen billones de células y menos de 30,000 genes ¿para qué, el código genético es más que un plan general? Por ejemplo, los vasos de sangres mayores se especifican genéticamente. Todos tenemos una aorta. Pero los vasos de sangre menores crecen donde necesitan según las reglas genéticamente definidas.
Aunque todas las células somáticas en un organismo contienen el código genético completo, en cualquier célula dada, sólo relativamente pocos genes son activos. La diferencia en los genes que son activos determina la diferencia entre, próstata, corazón y células del cerebro. La lógica del gen es compleja ya que determina cuando y donde un gen particular será “activado.”
La lógica del gen puede acomodar cantidades variantes de detalles posesiónales. El ojo, que tiene una estructura compleja en que las células adyacentes pueden ser muy diferentes probablemente exige a muchos genes llevar a cabo una estructura relativamente pequeña. El análisis molecular de genes de control del desarrollo del ojo, está proporcionando nuevas visiones sobre los procesos evolutivos. Un estudio sobre un rango de especies se examinó el gen Pax6, su expresión en el ojo hace pensar en una relación evolutiva más convergente que divergente. El análisis de otros genes de desarrollo, particularmente en el ratón y Drosophila, este pequeño díptero llamado mosca del vinagre (Drosophila melanogaster)-, plantean preguntas extensas sobre los mecanismos evolutivos. ¿Existe un origen evolutivo común para los ojos?
[xlvi]
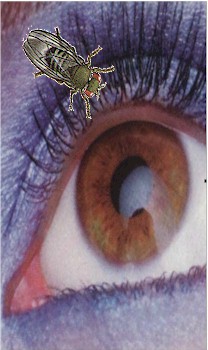
Fig. 2. Variedad de ojos.
Una variedad asombrosa de ojos existe a lo largo del reino animal, cada tipo es fino al ambiente, en donde un organismo particular vive. Esta diversidad de tipo de ojo sugiere que estas mismas estructuras diferentes, evolucionaron independientemente entre sí. De hecho el ojo tiende a menudo hacer usado como un ejemplo de independencia y de evolución convergente. Ha habido sin embargo, recientemente, mucha discusión de un linaje del ojo común, la evidencia tiene los genes reguladores, mismos que son compartidos entre los ojos tan diferente como aquéllos de Drosophila y ratón. Así los estudios de genes reguladores del desarrollo del ojo han adquirido una perspectiva evolutiva, y ha levantado la perspectiva de lo indefinible de esas asociaciones entre los organismos distantes. La evidencia acumulada para una fundamentación de la certeza de genes desarrolladores, es compartida por diversos linajes. Parece por consiguiente probable la existencia en el pasado de un órgano primitivo ancestral fotosensible, que requirió la presencia de estos genes
[xlvii]
.
El fémur es más grande que el ojo, pero mucho menos complejo y requiere de menos información genética. La lógica del gen también controla cuando varias actividades tienen lugar. Las células se dividen rápidamente en los organismos en formación pero no dividen en los adultos a menos que sea necesario reemplazar tejidos muertos o desechos celulares. El cáncer involucra una avería mayor en la lógica del gen, en que las células crecen en una posición impropia y "en un momento impropio”. Se piensa que el cáncer requiere mutaciones múltiples, algunas de las cuales pueden heredarse.
La lógica del gen usa las proteínas para llevar a cabo señalización. Es decir, los genes pueden controlar la producción de proteínas que realmente son los ladrillos para producir músculo y otros componentes estructurales en las células de organismos en formación, pero también pueden controlar la producción de otras proteínas que son los signos de la lógica. Éstos, los signos de la lógica pueden ser recibidos entonces por otros genes y determinan si esos genes se activan.
Algunas proteínas de la lógica son largas ya que en ellas pueden viajar a través del organismo entero –piense en la insulina-. Otros signos de rasgos más cortos sólo parecen cercanos a su punto de origen, posiblemente sólo inmediatamente alrededor de la célula en que ellos se generan. Los genes cortos y largos pueden generar y pueden descubrir las proteínas de señalización, muchos genes pueden llevar a cabo una lógica muy compleja. El armazón de lógica posicional que gobierna en el cuerpo, se encuentran en tipos específicos de células. El propio armazón de la lógica tiene que ser construida como un organismo que crece de una célula fertilizada, del huevo a un adulto.
Una mutación ocurre cuando el código genético en una célula se altera tal que esa célula descendiente también formada por la división de la célula alterada tiene el ADN alterado. Si la mutación ocurre en la cadena de división celular entre el huevo original fertilizado y reproductor (esperma o huevo) de las células (línea del germen), entonces la mutación puede pasarse a la descendencia. Muchas otras mutaciones no tienen efecto probablemente porque ellas ocurren en genes que nunca se activan en los descendientes de las células afectadas. (Una mutación en un gen que sólo es activo en el cerebro, no tendría efecto alguno si ocurriera en la línea de células que formarán una mano, etc.)
La evolución tiene lugar por medio de mutaciones que afectan la línea del germen. A menudo una mutación produce pérdida de alguna función esencial y es por consiguiente fatal a la descendencia y no pasa a los descendientes vivientes. A veces la mutación produce una ventaja evolutiva y por consiguiente puede colocarse como universal en el futuro en los descendientes. A veces la mutación produce características que son diferentes, como un color de ojos verdes en una especie que previamente tenía sólo ojos castaños, pero no confiere ninguna ventaja particular o desventaja y se pone común pero no universal en los descendientes. Los organismos superiores también tienen secciones extensas no-funcionales en su código genético. Las mutaciones en las partes no-funcionales del código no tendrían el efecto notable en el organismo y por consiguiente se pasarían a los descendientes. La proporción en que las mutaciones ocurren debe ser relativamente constante, pueden usarse diferencias en el código no-funcional para determinar el tiempo desde que dos individuos compartieron a un antepasado común.
El HGP indica que el genoma humano contiene 50 por ciento del código aparentemente no-funcional como muchos que consisten en muchas repeticiones de secuencias simples aproximadamente. ATATATATATAT…tiene un pequeño o ningún volumen de información. Algunas repeticiones se conocen como secuencias para ser modelos de sincronización, necesarios como las secuencias a los principios y extremos de cromosomas. El propósito, de cualquier otra repetición en las secuencias es desconocida.
En resumen, así que todo esto tiene que ver con el origen de la vida. El código genético representa un registro histórico del desarrollo del organismo con una cantidad extraordinaria de detalles (825 megabytes son muchos detalles). Un organismo que comparte las secuencias del código, significa muy probablemente que con otro organismo tiene un antepasado común. Mirando los cambios en el ADN no-funcional nosotros podemos estimar el tiempo desde que ese antepasado vivió. Comparando el genoma nosotros podemos construir un “árbol familiar” de vida en la Tierra.
Basado en los datos del HGP -Human Genome Project- reporta para el 23 de enero del 2003 un avance del 99.9%
[xlviii]
y apoyados en tras otras fuentes genómicas
[xlix]
podemos decir:
- Todos los humanos descienden de un solo individuo que vivió hace aproximadamente 270,000 años.
- Los humanos y los nuevos monos comparten un antepasado que vivió hace aproximadamente 7 millones de años.
- Los humanos y ratones comparten un antepasado común que vivió hace aproximadamente 50 millones de años.
- Se piensa que toda la vida en la tierra es descendida de un sólo organismo origen, de la célula primordial (primer criatura en el desarrollo) que vivió hace aproximadamente 3.5 mil millones de años.
- La Tierra fue formada hace aproximadamente 4.5 mil millones de años pero muy probablemente los 3.8 mil millones de años que datan los fósiles podrían ser relativamente incompatibles con la vida.
Cuando más datos del código genético estén disponibles sobre varios organismos, el análisis de diferencias y similitudes de progreso de los códigos en el árbol entero de la vida en la tierra, que se desarrollará en el futuro sin duda, más se sabrá sobre las características del organismo primordial. Quizás la respuesta de cómo la vida nació en la tierra pueda ser revelada, si es que ésta revela que nació en la tierra, tal como sostiene el renombrado astrónomo Fred Hoyle que apoya la teoría espacial.
[l]
2.4. Vino del espacio exterior
Algunos creen que la vida se originó en otra parte en el universo y se distribuyó entonces de algún modo. Esto no tiene que significar contaminación biológica de la Tierra temprana por viajeros espaciales que vacían sus tanques de basura. Basándose en evidencia de bacterias fosilizadas en los meteoritos y ADN que se ha recuperado de material que data de 20 millones de años de antigüedad. La posibilidad que la vida se allá originado en alguna otra parte en el universo (es un universo muy grande) y entonces vino a la tierra, parece la más probable idea. La teoría espacial también es menos egocéntrica. Tenga presente que toda creencia de “la Tierra como el centro del universo” han sido refutadas hasta hoy.
Una consecuencia de la teoría espacial considera que la vida podría distribuirse ampliamente. La vida podría aparecer relativamente rápida en cualquier planeta que tiene las condiciones apropiadas, por lo menos en regiones que estaban en una posición a ser sembrada la fuente. En otros términos, si hay vida en la Tierra, es probable entonces que allá vida en cualquier sistema cercano que tenga los planetas con las condiciones apropiadas.
¿Cómo, donde y cuando ocurrió el nacimiento de la vida? Hasta este siglo, es una clase de pregunta que normalmente se considera fuera del límite para la capacidad humana.
[li]
Dependiendo de su punto de vista, es para nosotros una señal de optimismo ilimitado que la ciencia y sus científicos hoy esperan resolver el enigma de cómo la vida empezó, lo decimos en lo absoluto sin ninguna arrogancia. En 1863, Charles Darwin comentó que era un ejercicio fútil -poco apreciado- para intentar aplicar el pensamiento científico a este origen de orígenes, cuando las primeras cosas vivientes asumieron y transformaron a nuestro planeta inanimado. Ocho años después él había cedido un poco, meditando más adelante, se preguntó si la vida podría haber empezado en algunos "estanque cálidos y pequeños" condimentados con especies químicas orgánicas simples.
¿Pero dónde en la Tierra podemos encontrar los ladrillos moleculares que formaron la vida, en un planeta que simplemente es una masa de piedra y agua? Nosotros nos hemos acostumbrado a la idea de una Tierra que germina la vida en cada nicho, que es difícil imaginarse el mundo yermo –inhabitado- de hace cuatro mil millones de años, cuando los mares fueron formados y el propio planeta era una mitad no más vieja -mil millones de años-. De algún modo, este mundo desovó las proteínas y los ácidos nucleicos -ADN y ARN- ésas son las huellas digitales moleculares que distinguen la vida.
Algunos científicos han especulado que estas moléculas no son en absoluto de cosecha propia, es decir esa vida se sembró del espacio, por esporas llevadas desde el profundo espacio helado a otra parte a través del vacío interestelar de un mundo viviente. Esta idea, se ha llamado "panspermia" en 1907 por el químico sueco Svante Arrhenius y se hizo revivir en los años sesenta por Francis Crick, el co-descubridor de la estructura del ADN.
Pero finalmente esto no sólo es satisfactorio como hipótesis, mientras ésta dirige hacia afuera la atención, la pregunta central la envía a otro lado, a otro lugar, pero sigue siendo científica -- porque no es obvio cómo pudiera probarse la vida-. La mayoría de los científicos prefiere asumir que las moléculas que constituyeron los organismos más primitivos surgieron de moléculas más simples, pequeñas formadas por procesos no biológicos en la Tierra primitiva.
¿Cuáles son estos elementales leíbles de la sustancia viva? Son las proteínas, largas cadenas de moléculas más pequeñas llamadas aminoácidos. Mucho del trabajo sobre el origen de vida se ha enfocado en la pregunta de cómo los aminoácidos se formaron y cómo ellos se unieron en proteínas. Los aminoácidos contienen carbono, hidrógeno, principalmente oxígeno y átomos de nitrógeno. Todos éstos elementos habrían estado presentes en alguna forma en la atmósfera de la joven Tierra: al contrario de la atmósfera de hoy, no era principalmente una mezcla de oxígeno y gas de nitrógeno, en cambio puede haber contenido el nitrógeno junto con monóxido de carbono o dióxido -emitido de volcanes-, o quizás el metano -un compuesto de carbono e hidrógeno-. Aunque las moléculas de aminoácidos son pequeñas y simples comparadas con las proteínas, ellas son complejas cuando se les compara con las moléculas de estos gases.
Pero el paso de las mezclas de gases crudos a los aminoácidos sofisticados no es tan complicado como podría parecer. En 1953, los químicos Urey y Miller’s de la Universidad de Chicago, mostraron que pueden hacerse unos aminoácidos simplemente mezclando amoníaco, hidrógeno, metano y vapor de agua en un frasco de vidrio y desintegrándolo con descargas eléctricas. Ellos sugirieron esto, dado que sería un poco como los relámpagos que existieron a través de los cielos prístinos –primitivos-.
Urey y Miller’s con su experimento dieron un giro, al persuadir a científicos que el origen químico de vida no es una desesperada propuesta. Pero realmente no proporciona ninguna respuesta firme. En primer lugar, el carbono en la atmósfera temprana estaba probablemente limitado a los óxidos del carbono, no en el metano. Si en cambio se usan los óxidos, la formación de aminoácidos en el experimento es despreciable. Otros esquemas, usando los materiales de arranque simples y las fuentes crudas de energías para estimular las reacciones, se han propuesto subsecuentemente como los mímicos de la manera que se podrían haber formado los aminoácidos; pero es justo decir que incluso este primer paso en busca del origen no resuelve hacia los restos de proteínas. Hay también una buena razón para creer que pueden formarse los aminoácidos en el espacio por reacciones que ocurren en las superficies de asteroides helados o meteoritos--ellos se han identificado, por ejemplo, en varios meteoritos ricos en carbono que se han colapsado en la Tierra-. Así aun cuando la vida no se puede desechar su origen el espacio, probablemente parece que algunos de sus ladrillos llegaron de esta manera.
Los eslabones de la cadena de ADN, entretanto, son más complicados. Ellos se llaman nucleótidos, compuestos de tres partes: una base, que se pega otra bajo una unión de doble hélice; una molécula de azúcar y un ion de fosfato. El fosfato esta en minerales -aunque es duro de formarlo en una forma soluble-. Desde que generalmente se cree que la química formativa de vida habría tenido lugar en el agua, éste ha sido un problema. Los azúcares pueden construirse de una molécula pequeña llamada formaldehído que podría haber estado posiblemente presente en la Tierra primitiva. Las bases de ADN son difíciles de sintetizar, pero nuevos químicos han descubierto las maneras creíbles de hacerlo en reacciones crudas, que involucran el cianuro de hidrógeno, una simple y pequeña molécula.
Por encima de todo esto la pregunta es cómo los ladrillos se unieron en las cadenas. Esto es lo más problemático que podría haber aparecido al principio. En primer lugar, el agua tiene una tendencia a dividir el eslabón, es decir separar a los aminoácidos. Una manera prometedora alrededor de esto es suponer que la vinculación pasó en las superficies de minerales donde los aminoácidos podrían ligarse. Químicos han mostrado que ciertos minerales comunes, como un tipo de arcilla llamados ILLITE -minerales de arcilla que tienen la estructura de cristal de muscovite esencialmente-, pueden catalizar la unión de aminoácidos. Otro de arcilla, montmorillonite -un mineral arcilloso suave que es un hidroxilo aluminio silicato con la capacidad considerable de intercambiar parte del aluminio por bases y magnesio, puede ayudar a la formación de cadenas de nucleótidos.
[lii]
La razón detrás de todos estos esquemas es que si los mares fueron finamente un combinado de todas las moléculas pequeñas en cantidades diminutas formadas de los constituyentes básicos de la atmósfera, entonces ellas se podrían haber concentrado en ambientes cálidos, mientras las albuferas -almacenamiento de líquidos- costeras se evaporan y conspiran para combinar en un rociar aún más fino los componentes de proteínas y ADN. Todo requiere un salto considerable de fe, pero el punto es mostrar que la vida sólo podría haber empezado de una manera parecida en cualquier lugar del universo, requiriendo algo extraordinario como una “piel arrojada de la ventana de alguna nave espacial visitante”. Se ha investigado el origen de la vida a partir de la naturaleza que gobierna está, que por cierto esta por todas partes, nuca debemos establecer la plausibilidad, mientras no se encuentren las pruebas.
Otro pensamiento, llamado escolar es un guión muy diferente, sin embargo, propone: apunta a los ecosistemas que crecen alrededor de las primaveras calientes llamadas ventilas hidrotérmicas en el suelo de los océanos profundos, fuera de alcance de la luz del sol que finalmente apoya a la mayoría de las comunidades vivientes. En las comunidades de la ventila hay organismos que pueden vivir del calor y de beber ricos nutrientes minerales y gases que vierten estas aberturas, les gusta el humo espeso de las chimeneas de la abertura volcánica. ¿Quizás las aberturas hidrotérmicas proporcionaron ambos materiales crudos de la vida primitiva y la energía necesaria para conseguir que ellos reaccionaran para hacer las moléculas más complejas? Es una idea contenciosa; algunos expertos, como Miller Stanley, contienden que las aberturas harían más para quemar a las moléculas complejas que para crearlas.
Pero aun cuando nosotros podemos deducir cómo hacer los componentes de las proteínas y los ácidos nucleicos y atarlos juntos, nosotros no habremos obtenido una receta de la vida. Las proteínas no son el azar de probabilidad cero, no son sólo ataduras de aminoácidos -ellas incluidas forman información puesta en código en la secuencia específica en que los aminoácidos se unen-. Esta secuencia determina la forma de la cadena plegada de la proteína que a su vez determina su función biológica. La información para una proteína es puesta en código en los genes del ADN, en la hélice doble. Pero para traducir y copiar esta información, el ADN necesita la ayuda de proteínas. Así que el rompecabezas central es: ¿qué vino primero, ADN o proteínas?
La conclusión actualmente no es una cuestión de estar a favor o en contra. En cambio, investigadores creen, que la "vida" más primitiva puede haber contado con un ADN primo originado del RNA. Es del agrado de muchos considerar el ADN un caso particular de la RNA. La ARN puede poner en código las instrucciones para una proteína en su secuencia de nucleótidos; de hecho, una secuencia de ADN se copia primero en la forma de ARN antes de que se tradujera en una proteína en las células. Pero ARN tiene otro talento: en ocasiones puede actuar como una proteína catalizadora de las reacciones químicas. El descubrimiento de "ARN catalizador" en los años ochenta por los químicos premio Nobel 1989, Sidney Altman y Thomas Cech empujaron la idea que la vida más primitiva era un "mundo de ARN" en que el ARN hizo todo el trabajo duro, sólo para ser relegado finalmente al mensajero del ADN, una vez formadas las proteínas constitutivas de la célula
[liii]
.
Cuestión de Oxigeno. La vida tal vez vino de oscuros, húmedos y aparentemente malolientes lugares naturales en donde de los más extraños eucariotes no se sospechaba su presencia. Gustaban de zonas pantanosas, para pulular principalmente estas células simples eucarióticas –protistas-, tal como todas las células, estas deben producir ATP para sobrevivir. Estos lugares todavía no contienen suficiente oxígeno para sostener la síntesis de ATP.
[liv]
Algunos protistas no poseen mitocondria, sobreviven de la fermentación anaerobia en el citosol. Por ejemplo el ciliado vive sofocantemente en los intestinos pobres de oxigeno de la cucaracha dónde ayuda a que el insecto digiera la celulosa. En lugar de consumir oxigeno el mitocondrion Nyctotherus tiene la propiedad extraña de excretar el hidrógeno como un derivado de la síntesis de ATP. Organelos generadores de hidrógeno similares -hidrogenosomas- se han estudiado en eucariotes anaerobios por 25 años.
[lv]
Hidrogenosomas han sido a menudo sospechosos de provenir de la misma bacteria propuesta por la teoría endosimbiótica –origen de las mitocondrias actuales-. Pero Akmanova informa que un hidrogenosoma tiene su propio genoma, mostrando con esto su pasado endosimbiótico directamente
[lvi]
,
[lvii]
. Células Nyctotherus qué no crecen en cultivos y tienen que ser manejadas cuidadosamente, es decir micromanipuladas en los intestinos posteriores de la cucaracha encierran hidrogenosomas que pueden estar etiquetados por los anticuerpos en contraste con el ADN.
Investigadores encuentran que la célula produce un ribosomal ARN que aunque no ha demostrado por hibridación en situ localizado en el organelo, llevan toda la secuencia característica esperada para mitocondrias ciliate. Más puede parecer a un mitocondrion, excepto que este orgánulo DNA, es indiscutiblemente un hidrogenosoma porque produce hidrógeno. Akmanova encontró consumo de hidrógeno metanogénico endosimbiótico dentro de las células de Nyctotherus. Finalmente el Nyctotherus expresa un gen de codificación nuclear para un hidrogenosoma -una enzima que hace el hidrógeno- eso probablemente se importa en el hidrogenosoma con un transporte péptido.40
La importancia evolutiva de estos resultados es doble, primero en la asociación DNA hidrogenosoma era hasta aquí un genuino eslabón perdido
[lviii]
. La reducción de nitrato NO3 a nitrógeno gaseoso N2 se denomina desnitrificación, estas bacterias son anaerobias (Nyctotherus).


Fig. 3. Protozoario Ciliado dentro del intestinos de un Discus.
Nyctotherus. Este organismo se encontró en el intestino de un Discus –Symphysodon, Fig. 3- del Sudeste de Asia. La apariencia vacuola contráctil, el macronucleus y el posible micronucleus, es similar a especies de Nyctotherus que se encuentran en el colon de anfibios y en algunos invertebrados como las cucarachas.
El biólogo evolucionista "estudia los pasos de las adaptaciones milagrosas tan características de cada aspecto del mundo orgánico evolucionado"
[lix]
. Pero la naturaleza general de tales pasos adaptables todavía es incierta. Se piensa a menudo que la evolución es producto de eventos imprevisibles –aleatorios-. Podríamos en este mismo sentido esperar que los pasos tomados por la adaptación deban ser aleatorios, biológicamente y temporalmente.
Con ayuda de la teoría de derivación matemática (series de tiempo) se muestra que los pasos adaptables pueden tener un ritmo bastante fuerte. Explica que la fuerza del ritmo adaptable es su regularidad temporal relativa, es igual a una constante, misma para toda población microbiana
[lx]
. Como una consecuencia, se predicen números de adaptaciones acumuladas para tener una proporción de la varianza/media índice de variación. La teoría derivada, es potencialmente aplicable al estudio de evolución molecular. Las poblaciones de organismos se adaptan a su ambiente a través de la producción de mutaciones beneficiosas y la fijación subsiguiente de estas mutaciones al predominio en la población a través de la selección natural, un proceso conocido como fijación. La fijación de una mutación beneficiosa dada, toma a menudo un gran tiempo. El resultado es la presencia simultánea en la población de varios nuevos linajes que cada uno lleva una ventaja selectiva encima de su progenitor común. En las poblaciones microbianas, la unión genética (es decir, escasez de recombinación) entre las mutaciones beneficiosas, en un fragmento grande de estos linajes causará permanecer en la competición directa a lo largo del concurso resultante para la fijación. Este período de competición introduce un grado de previsibilidad por el tiempo que pasa antes de una ganancia, o éxito, el linaje es fijo. El resultado es la uniformidad relativa en los intervalos de tiempo entre las fijaciones.
El fenómeno de competición entre linajes creados por las mutaciones beneficiosas se ha llamado "efecto Hill-Robertson" para la población sexual
[lxi]
e "interferencia clonal" para la asexual.
[lxii]
Los efectos fueron deducidos por Fisher y Muller, históricamente, este fenómeno ha sido considerado más a menudo en el contexto de discusión y modelos de la ventaja evolutiva de sexo: puede verse como una fuente de ineficacia en selección natural que puede remediarse por la recombinación aumentada. Los datos de los experimentos con el coli Escherichia y el virus stomatitis vesicular, confirmaron la predicción, que, como proporción de la mutación o aumentos de tamaño de población, el número de competidores de los linajes debe aumentar y por consiguiente (1) la ventaja de aptitud del linaje premiado debe aumentar y (2) la proporción en que los linajes premiados están fijos debe aumentar a medida que disminuyen la velocidad. Cabe aclarar que no se dirigió la regularidad temporal de adaptación, sin embargo, en los experimentos o los modelos anteriores Gösta Eriksson60 se enfoca en el aspecto temporal de adaptación. Su simplicidad relativa revela su generalidad. Muchas mutaciones beneficiosas existen breve tiempo, estas mutaciones casi nunca logran una alta frecuencia para afectar la aptitud de la población y son así inconsecuente de un punto de vista evolutivo. Además, una mutación beneficiosa puede ocurrir en un genoma que contiene uno o las mutaciones más deletéreas. Si la aptitud neta del linaje mutante resultante es más bajo que la población, entonces esta mutación beneficiosa ciertamente se perderá.
Las mutaciones beneficiosas de interés son sólo aquéllas que sobreviven la tendencia genética y no se unen a las mutaciones fuertemente deletéreas.
[lxiii]
La evolución rápida de genes reproductores masculinos en el linaje del hombre también es tema de estudio, ¿la velocidad es importante de terminarla?
¿Hace cuánto que aparecieron los primeros vertebrados? Un fósil con 350 millón de años de antigüedad, proporciona evidencia de una fase casi desconocida en el origen de vertebrados de la tierra. También es un recordatorio del poco conocimiento de las relaciones entre los linajes principales de anfibios y reptiles. La transición entre los peces y vertebrados de la tierra era un punto que regreso con este hallazgo de la historia de la vida. El fósil, Pederpes finneyae (fig. 4), viene de los depósitos Dumbarton:


Fig. 4. Pederpes finneyae.
El espécimen fósil encontrado en 1971, el pie de la parte de atrás tiene una estructura que ayuda la andadura
[lxiv]
se reprodujo casi ciertamente en el agua, un poco como las salamandras acuáticas modernas Pederpes finneyae.
[lxv]
2.7. Evolución y origen: Código C-3
El concepto de evolución fundamentalmente no sólo está dentro de la teoría biológica como una idea sin aplicación, también encuentra un rico y profundo espacio de aplicaciones en la bioingeniería y biotecnología. En particular la evolución invitro se ha usado ampliamente en el estudio de la evolución molecular temprana, es decir, en modelos sobre el origen de los sistemas genéticos, por ejemplo, el cómo las células procesan la información del ADN.
[lxvi]
La aparente distribución no aleatoria de la asignación de aminoácidos dentro de un código genético, provoca las preguntas siguientes
[lxvii]
: ¿nos dice algo la estructura del código sobre la evolución?, y ¿cuál podría ser el algoritmo que la gobierna? El código genético podría ser un accidente histórico que fue estable en el último antepasado común de los organismos modernos. Los argumentos de adaptabilidad, históricos y químicos, sin embargo, desafían a semejante modelo de accidente. Estos argumentos proponen que el código actual es de algún modo óptimo, refleja la expansión de un código más primitivo que permitió incluir más aminoácidos, o es una consecuencia de interacciones químicas directas entre ARN y aminoácidos, respectivamente. Tales modelos son mutuamente excluyentes, sin embargo, ellos pueden reconciliarse con un modelo estereoquímico evolutivo de interacciones que formaron el código inicial que como consecuencia se extendió a través de la transformación biosintética de aminoácidos codificados y, finalmente, se perfeccionó a través de la reasignación del codón. Alternativamente, podrían haber actuado para asignar los 20 aminoácidos en armonía -los aminoácidos naturales- a sus posiciones presentes en el código genético.
Una teoría cuantitativa de un proceso evolutivo requeriría una comprensión cuantitativa del proceso de selección, como también del modelo evolutivo llamado sembrado-medio para la evolución de ARN en poblaciones de virus. La teoría opera con una distribución de la población en un espacio unidimensional.
[lxviii]
Basado en los experimentos evolutivos de ARN-virus se deriva un modelo en procesos evolutivos para un individuo, se da como la suma de muchas contribuciones individuales que pueden deformarse independientemente. Los estudios demostraron una relación fuerte entre la teoría y las simulaciones para tamaños de poblaciones pequeñas y para el equilibrio, pero la dinámica de la evolución resultó ser patológica en los tamaños de poblaciones grandes donde se amplifican las mutaciones exponencialmente -sumamente raras-, reproducción evolutiva a velocidad infinita.63
La teoría de coevolución
[lxix]
,
[lxx]
define a un aminoácido precursor, como uno en el que cualquier proporción del aminoácido está incorporado metabolicamente dentro de un producto aminoácido. El producto aminoácido es definido como el aminoácido que mantiene un menor número de pasos metabólicos desde el aminoácido precursor.
[lxxi]
El metabolismo aminoácido es muy complejo porque no sólo sirve para producir los 20 aminoácidos usados por la biosíntesis de proteína, además funciones de señalización como una fase para la síntesis de muchas moléculas importantes y raros aminoácidos (por ejemplo la ornitina) nos indican la amplitud de sus funciones.
[lxxii]
Tabla 1. Pares producto-precursor63
|
Glu
 Arg
Arg |
Asp
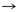 Asn
Asn |
Ser
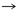 Trp
Trp |
Thr
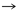 Ile
Ile |
Val
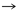 Leu
Leu |
Glu
 Gln
Gln |
Asp
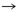 Thr
Thr |
Ser
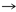 Cys
Cys |
Thr
 Met
Met |
|
Glu
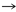 Pro
Pro |
Asp
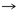 Lys
Lys |
Phe
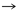 Tyr
Tyr |
Gln
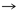 His
His |
|
Asp
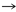 Arg
Arg |
Asp
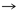 Asn
Asn |
Ser
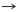 Trp
Trp |
Thr
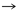 Ile
Ile |
|
Glu
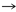 Gln
Gln |
Asp
 Thr
Thr |
Ser
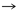 Cys
Cys |
Asp
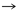 Met
Met |
|
Glu
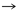 Pro
Pro |
Asp
 Lys
Lys |
Phe
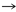 Tyr
Tyr |
Gln
 His
His |
|
|
De acuerdo con la tabla 1, 13 pares de producto-precursor definidos por coevolución se muestran en las primeras tres líneas. La lista de 12 pares fundada en una definición bioquímica creíble, par precursor-producto se muestra en las últimas tres líneas. Las diferencias con la teoría de la coevolución están en negrita.
[lxxiii]
|
Se han apoyado los científicos en la estadística para abordar la coevolución, se aplica la distribución hipergeométrica a cada par producto-precursor para calcular la probabilidad que habría para un sólo caso sobre el número observado de codones asignados a los aminoácidos del producto para quedar en una sola mutación fuera del punto de los codones precursores, tal como lo indica la siguiente expresión matemática.

donde “c” denota el número total de codones que quedan en un punto de mutación de los codones del aminoácido precursor, “g” denota el número de codones que quedan en más de un punto de mutación del codón del aminoácido precursor, “w” denota el número de codones del producto que queda sólo en un punto de mutación de los codones del precursor (es decir, ese encaja la predicción de coevolución), y “n” denota el número total de codones del producto. En otros términos, esta ecuación evalúa la pregunta combinatoria: dadas las asignaciones de un aminoácido precursor dentro del código, ¿cuál es la probabilidad de asignación aleatoria de “n” codones del producto aminoácido que produciría “w” o más ajustable a las predicciones de la teoría coevolucionaría, es decir, de que quede dentro un punto de mutación por lo menos un codón del precursor.
Mediante técnicas de experimentación computacional a nivel molecular se demuestra la viabilidad de confrontar este modelo y otros que podrían arrojar su correspondencia con respecto al comportamiento de la naturaleza de la vida.
[lxxiv]
Podríamos decir que la evolución de proteínas se codificó en secuencias de nucleótidos para iniciar con el avenimiento de un código de longitud tres, C-3. El orden cronológico de la aparición de aminoácidos sobre el escenario de la evolución y los pasos en la evolución en la reconstrucción del C-3, el investigador Trifonov los aborda de una manera muy original.
[lxxv]
Trifonov reconstruye con 44 criterios e hipótesis cronológicas para los aminoácidos – ver Tabla 2-. Según la cronología general, el par complementario codón GGC y CCG del aminoácido Alanina y Glicina se presume aparecieron primero en la evolución. Otros codones aparecen como pares complementarios que dividen su respectivo aminoácido en dos alfabetos, codificados por tripletas de purinas y pirimidinas centrales: G, D, S, E, N, R, K, Q, C, H, Y, y W (el alfabeto de Glicina G) y A, V, P, S, L, T, I, F, y M (Alanina alfabeto A). Se especula que las cadenas tempranas de polipéptidos eran muy cortas, probablemente de una longitud uniforme, perteneciendo a dos tipos de alfabetos de codificación, en dos cuerdas dúplex ARNm tempranas. Después de la fusión de los mini-genes, un mosaico de alfabetos se formó, el trazo de la estructura, implica detectar en las proteínas secuencias de genomas procariotes completos en la formación de oscilaciones débiles con periodos de 12 productos en la forma de alteración de dos tipos de 6 unidades de productos largos.
Tabla 2. Cuarenta y cuatro criterios e hipótesis para la cronología de aminoácidos.

Fig. 5. El mapa secuencial de la evolución del código genético basado en el par precursor-producto dentro de la teoría de coevolución.
Los recuadros mezclados de la figura 5, indican que las asignaciones de los codones primordiales putativos, exigieron crear las relaciones por teoría de coevolución. Los codones puestos en letra itálica no concuerdan con las predicciones de coevolución. Proponiendo a nuestro intelecto una teoría biosintética del código genético: pregúntese, lo que vemos es ¿verdad o ficción?63.
Un argumento frecuente en apoyo a la idea sobre la probabilidad de vida, es una estimación de probabilidad de originarse por causalidad, una secuencia de proteína significante, un polipéptido de unos 100 aminoácidos. El número de diferentes posibles moléculas de esta longitud está en el orden de 10x130, un tamaño muy grande, es decir sobre el número de átomos en el universo visible. Esta estimación es sin embargo un tanto desconcertante. Ciertamente desde que la vida empezó, eran moléculas muy pequeñas, por ejemplo, la oxitocina producida por células neurosecretorias en el hipotálamo son péptidos cortos de nueve aminoácidos –una hormona-. La actividad péptida biológicamente más frecuente tiene el tamaño de 20 a 40 productos y se conoce las funciones de mini-genes, incluyendo 2 codones. La evidencia de tales fases simples es proporcional al tiempo inicial por análisis de autocorrección de secuencias de proteínas de procariotes. Así la idea de coevolución se ha establecido para describir varias fases distintas en la evolución en proteínas nucleicas en código, desde lo simple a lo complejo. Reflejándose ambos extremos en la estructura de proteínas modernas y sus secuencias. Las fases o etapas sugeridas por Trifonov75:
I.- Homopeptidos cortos Glin y Alan.
II.- Secuencias mixtas de 6 residuos-largos péptidos de alfabetos de Glicina (G) y Alanina (A).
III.- de 25-30 residuos-largos, péptidos cerrados dentro de la vuelta por los contactos extremo a extremo.
IV.- 100-200 residuos-largos, proteínas plegadas (dominios)
V.- Multidominio proteico. Gen que probablemente entra en las fases o etapas IV-V de semejante manera las regularidades del tamaño más tempranas todavía se respetan.
[lxxvi]

Fig. 6. Las fases o etapas sugeridas por Trifonov.
El rango de aminoácidos en proteínas primitivas fue presumiblemente muy limitado. Una pregunta natural es: ¿Qué aminoácidos fueron los primeros en aparecer y en qué orden cronológico se efectuaron todos los otros aminoácidos que aparecieron? Cada una de las numerosas teorías del origen del código genético, y varias otras consideraciones, sugieren que los aminoácidos tienen un cierto orden temporal. Otros se apoyan en teorías que consideran lo contrario. Uno podría pensar en un acuerdo equilibrado que tendría en cuenta las distintas estimaciones. Sin embargo, la pregunta de peso para poder dar un criterio de cómo surge, hace que cada esfuerzo de este tipo inevitablemente sea subjetivo y dudoso.
Una manera justa y razonable de derivar el acuerdo general es por promedio de líneas diferentes, sin pesos dados, excepto por eliminación de aquellas que son casi idénticas mientras se combinan así en un criterio. Semejante análisis ya mencionamos que se realiza de hecho con 40 diferentes criterios de cronología de aminoácidos por Trifonov. Entre criterios y teorías esta nos hace pensar la existencia de un orden específico dentro de los aminoácidos y los codones, como la ya mencionada teoría de Coevolución de Wong, la teoría RNA de Eigen, teoría Jukes y otras hipótesis
[lxxvii]
.
El criterio basado en simplicidad química de aminoácidos, en su reactividad, o en la composición de proteínas tempranas también fue incluido. Teniendo todo en consideración, un notable orden en ambas cronologías referentes a aminoácidos o codones, es revelada:
Los experimentos de imitación por Miller(1953-1987) están en primer lugar.
[lxxviii]
Además de los experimentos de condiciones interestelares para las condiciones prebióticas.
[lxxix]
Los codones parecen haber estado comprometidos por pares complementarios.
[lxxx]
Los pares de codones más estables están comprometidos primeramente.
Los nuevos codones simplemente son derivados del punto de cambio de los previos primeramente comprometidos.
Una visión moderna de la cronología de aminoácidos y codones es representada en la tabla 2 y figura 5. Estos son el resultado de los cálculos realizados del trabajo citado de Trifonov. La condición de criterios de cronología es basada en la reconstrucción de la composición de aminoácidos del antiguo ferredoxin.
[lxxxi]
Tabla 3. Cálculo del consenso cronológico de aminoácidos sobre los 36 criterios básicos de Trifonov.
|
Alineación
media |
Error
|
orden |
G |
4.7 |
0.8 |
1 |
A |
5.2 |
0.9 |
2 |
V |
6.8 |
0.7 |
3 |
D |
7.3 |
0.7 |
4 |
S |
8.0 |
0.7 |
5 |
E |
8.5 |
0.7 |
6 |
P |
8.8 |
0.8 |
7 |
L |
9.7 |
0.8 |
8 |
T |
10.2 |
0.6 |
9 |
N |
11.5 |
0.7 |
10 |
R |
11.5 |
0.7 |
11 |
I |
11.6 |
0.7 |
12 |
K |
11.8 |
0.8 |
13 |
Q |
11.9 |
0.7 |
14 |
C |
12.3 |
0.8 |
15 |
F |
12.3 |
0.8 |
16 |
H |
13.1 |
0.7 |
17 |
M |
14.3 |
0.6 |
18 |
Y |
14.4 |
0.6 |
19 |
W |
15.8 |
0.6 |
20 |
Las estimaciones de alineación promedio para la mayoría de los aminoácidos permiten ser ordenados singularmente en la cronología. Notablemente los aminoácidos de Miller’s en su mezcla están dentro de la exactitud de las estimaciones. Esto significa primeramente que la vida emerge de todos los aminoácidos que ya estaban presentes en el ambiente, es la evidencia fundamental de la naturaleza temprana y sus procesos de vida.
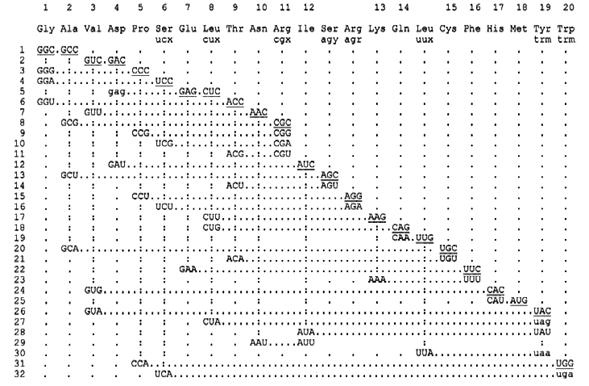
Fig. 7. Reconstrucción de la cronología codón.
La figura 6 presenta la reconstrucción para el orden temporal de los codones, basado sobre la discusión cronológica de los aminoácidos, y sobre la original sugerencia de Eigen y Schuster, sobre la primacía de la termoestabilidad y complementaridad (los números superiores corresponden al alfabeto: G A V D S E P L T N R I K Q C F H M Y W).
[lxxxii]
,
[lxxxiii]
Esta figura 7 es una versión modificada del esquema de Trifonov. El Orden de los aminoácidos P, S y E es ligeramente cambiado para empatar con los errores de la columna en la tabla 3. Las líneas del esquema corresponden a los pares de codones complementarios en cada arreglo de codones (conjuntos verticales) la mayoría de codones estables (subrayados) ocupan las más altas posiciones. Un rasgo que llama la atención de la figura 6 es el arreglo diagonal bajo, de los codones respectivos. Cualquier cambio sustancial en el orden de los aminoácidos (alineación superior) habría destruido el módulo global triangular. El patrón triangular significa que el consenso cronológico de los aminoácidos y sus roles termoestables y complementarios juntos, dictan una regla más natural del rol de la existencia de los primeros como un sólo punto de mutación –un rol procesal-.
La mayoría de los nuevos codones son generados por mutaciones y copiado complementario, por lo contrario el par GUC/GAC para V y D se derivan de la transición putativa en las posiciones medias de los codones primitivos GGC y GCC. El primer par codón complementario GGC*GCC, deben corresponder a los códigos primitivos del ARN duplex, con la cuerda complementaria GGCGGCGGC…GGC y GCCGCCGCC…GCC de cierta longitud que codifica para GlyGlyGly…Gly y AlaAlaAla..Ala, respectivamente. La composición original aminoácida de las proteínas es de 50% de Gly y 50% de Ala. En las fases posteriores de la evolución de la proteína la abundancia de Gly y Ala fue reducida gradualmente por otros aminoácidos, reduciéndose posteriormente a valores actuales del 6% y 8% respectivamente. El papel evolutivo excepcional de la Gly es ilustrado por la siguiente observación importante. La caída en el contenido de Gly es observada verdaderamente cuando las secuencias de proteínas funcionalmente relacionadas en procariotes y eucariotes son alineadas y la composición de partes comunes de las secuencias es calculado por Trifonov. La proporción de la Gly en los residuos compartidos es tan alta como 14%.
[lxxxiv]
Esta evaluación de los contenidos de la Gly es probable que corresponda al momento de separación entre eukriotes y procariotes, cerca del 3.5 mil millones de años -Doolittle, citado por Trifonov-.
[lxxxv]
Así el orden de Gly puede servir como un reloj para construir el árbol evolutivo. Recientemente un árbol mayor a seis reinos es presentado por Trifonov.82
Ver la serie de diapositivas expuestas por
 :
:
http://dieumsnh.qfb.umich.mx/codigo3/
Ahora podemos recurrir al principio cronológico por coevolución, explicar cómo algunos aminoácidos surgen de los de longitud más cortos, estos últimos asociados a los primordiales. Bajo poco más de 40 criterios formados por hipótesis y principios, Trifonov nos ayudó recrea un modelo de coevolución en donde sólo los pares producto-precursor Asp
 Met y Asp
Met y Asp
 Arg no aparecen en las pruebas de simulación computacional. Además es claro que el número de intercambios de palabras código entre un aminoácido –secuencia de palabras C-3- y otro, nos proporciona que aminoácido es precursor de cual. Considerando todo esto, nosotros especulamos que un sistema biológico óptimo reprime la recombinación genética cuando su genoma es óptimo –proceso que busca la adaptación óptima a cambios del medio ambiente-. Esto nos lleva a la idea de un algoritmo recursivo de instrucciones de recombinación, donde la condición de optimización rompe el programa cuando la secuencia del genoma es óptima para el sistema, reprimiendo el algoritmo de mutaciones sólo hasta cuando el sistema biológico ya no es óptimo frente al contexto en el que se mueve.
[lxxxvi]
Arg no aparecen en las pruebas de simulación computacional. Además es claro que el número de intercambios de palabras código entre un aminoácido –secuencia de palabras C-3- y otro, nos proporciona que aminoácido es precursor de cual. Considerando todo esto, nosotros especulamos que un sistema biológico óptimo reprime la recombinación genética cuando su genoma es óptimo –proceso que busca la adaptación óptima a cambios del medio ambiente-. Esto nos lleva a la idea de un algoritmo recursivo de instrucciones de recombinación, donde la condición de optimización rompe el programa cuando la secuencia del genoma es óptima para el sistema, reprimiendo el algoritmo de mutaciones sólo hasta cuando el sistema biológico ya no es óptimo frente al contexto en el que se mueve.
[lxxxvi]
La teoría de la evolución es un principio que habla de un intercambio dentro de una diversidad genética, haciendo de lado la información de la vida en sociedad de las especies. Señalamos con ello, que la evolución debe considerar además el intercambio de información en la vida social de las especies, dado que esta se traduce en cambios fisiológicos. Hablar de coevolución genético-cultural es agregar una variable más al problema de la evolución, pero muy necesaria. Se han desarrollado modelos de evolución por culturalización, evidencia que se muestra en el reporte de investigación del científico Kevin N. Landa de la Universidad de Cambridge.
[lxxxvii]
¿Cómo leer el código C-3?
El C-3 puede caracterizarse por los codones de ARNm, hay 64 codones posibles, la columna de la izquierda de la tabla 4 contiene la primera letra de los codones. En la primera fila esta la segunda letra y la columna de la derecha la tercera letra. Si requiere saber que aminoácido está codificado por GAU, se debe buscar la primera letra en la columna de la izquierda “G”, la segunda en la fila uno “A” y la tercera en la columna de la derecha “U”. En donde se interceptan las tres está indicado el aminoácido Asp –ácido aspártico-. Se llaman codones sinónimos los que codifican para el mismo aminoácido.
Tabla 4. C-3
|
U |
C |
A |
G |
|
U |
UUU
UUC
UUA
UUG |
Fen
Leu |
UCU
UCC
UCA
UCG |
Ser |
UAU
UAC
UAA
UAG |
Tir
Paro
Paro |
UGU
UGC
UGA
UGG |
Cis
paro
Tri |
U
C
A
G |
C |
CUU
CUC
CUA
CUG |
Leu |
CCU
CCC
CCA
CCG |
Pro |
CAU
CAC
CAA
CAG |
His
Gln |
CGU
CGC
CGA
CGG |
Arg |
U
C
A
G |
A |
AUU
AUC
AUA
AUG
|
Ile
Met
inicio |
ACU
ACC
ACA
ACG |
Tre |
AAU
AAC
AAA
AAG |
Asn
Lis |
AGU
AGC
AGA
AGG |
Ser
Arg |
U
C
A
G |
G |
GUU
GUG
GUA
GUC |
Val |
GCU
GCC
GCA
GCG |
Ala |
GAU
GAC
GAA
GAG |
Asp
Glu |
GGU
GGC
GGA
GGG |
Gli |
U
C
A
G |
Constrúyase un ensayo corto agrupando las teorías de coevolución química, evidencia fósil y criterios para el árbol de la vida; reportándolo en el formato de comunicación bajo alguna norma de estilo editorial y cuidando que las fuentes de información y la terminología de las proposiciones derivadas que respeten la complejidad original. Prepare una exposición argumentativa para defender su trabajo, bajo el esquema intelectual llamado discusión.
Abiogénesis, Ala, Aminoácido, Antrópico, Árbol de la vida, Arg, Arqueobacteria, Asn, Asp, ATP, Bacteria, Bases nucleotidas, Bioingeniería, Biosíntesis, Cáncer, Cianobacteria, Cianotipo, Cis, Código, Codón, Coevolución química, Cromosoma, Densidad de probabilidad, Distribución de probabilidad, DNA, Endosimbiótica, Eón, Estromatolitos, Eukarya, Fen, Fenotipo, Fuerza del ritmo, Gen, Genealogía, Genoma, Gli, Gln, Glu, HGP, Hidrogenosoma, His, Ile, Insulina, Interferencia clonal, Isótopo, Leu, Línea de germen, Lis, Lógica genética, Met, Meteorito, Microfósil, Mitocondria, Mitocondria, Modelo Bayesiano, Modelo de accidente, Mundo yermo, Mutación, Nucleótidos, Nyctotherus, Organismos primordial, Orn, Panspermia, Pax6, Pericentrómeros, Polipéptido, Precámbrico, Pro, Pro, Productos aminoácidos, Proteína, Química prebiótica, rARN, Reloj molecular, Ribosoma, RNA, Ser, Sopa prebiótica, Telómeros, Teoría espacial, Tierra prebiótica, Transposons, Tre, Tri, Val, Variabilidad Genética.
[vi]
Nakamura Y, et al. Complete genome structure of Gloeobacter violaceus PCC 7421, a cyanobacterium that lacks thylakoids. DNA Res. 2003
Aug 31;10(4):137-45. Recuperado el 13 de abril de 2010, de http://www.ncbi.nlm.nih.gov/pubmed/14621292
[x]
Larry S. Liebovitch, Yi Tao, Angelo T. Todorov, and Leo Levine. Is There an Error Correcting Code in the Base Sequence in DNA? Biophysical
Journal 71:1539-1544 (1996). Recuperado el 12 de abril de 2010, de http://www.ccs.fau.edu/~liebovitch/B60067EB.PDF
[xiii]
S. J. Mojzsis, G. Arrhenius, K. D. McKeegan, T. M. Harrison, A. P. Nutman and C. R. L. Friend,
1996, Evidence for life on Earth before 3,800 million years ago. Nature, 384, 55-59. Recuperado el 13 de abril de 2010, de
http://www.nature.com/nature/journal/v384/n6604/abs/384055a0.html
[xiv]
Jaynes, E. T., 1979, Where do we Stand on Maximum Entropy?, in The Maximum Entropy
Formalism, R. D. Levine and M. Tribus (eds.), M. I. T. Press, Cambridge, MA. Recuperado el 13 de abril de 2010, de
http://bayes.wustl.edu/etj/articles/
[xvii]
Higgs PG, Pudritz RE. (2009) A thermodynamic basis for prebiotic amino acid synthesis and the nature of the first genetic code.
Astrobiology. 2009 Jun;9(5):483-90. Recuperado el 16 de junio de 2010, de http://www.ncbi.nlm.nih.gov/pubmed/19566427
[xxxiii]
D. R. Bentley, P. Deloukas, at. alt., The physical maps for sequencing human chromosomes 1, 6, 9, 10, 13, 20 and X, Nature 409, 942 - 943
(2001) Recuperado 16 de junio de 2010, de http://www.nature.com/nature/journal/v409/n6822/full/409942a0.html
[xl]
San Miguel, P. , Gaut, B. S. , Tikhonov, A. , Nakajima, Y. & Bennetzen, J. L. Nature Genet. 20, 43–45 (1998)
[lii]
Sandrine Demaneche et al. Evolution of Biological and physical protection agaienst nuclease degradation of Clay-Bound plasmid
DNA. A and E. Microbiology, 67(1): 293-299 (2001) 8 Recuperado 16 de junio de 2010, de http://aem.asm.org/cgi/content/full/67/1/293
[lvi]
Hector l. Balbarrey, Juan C. Picena, Carlos A. Poy, Edgardo E. Quibert, Apoptosis e insuficiencia cardíaca,Rev Fed Arg
Cardiol 2001; 30: 71-77, Recuperado 5 de julio de 2010, de http://www.fac.org.ar/faces/publica/revista/01v30n1/balba/balba.PDF
[lix]
John Wilkins, Evolution and Philosophy Naturalism: Is it necessary? (1997) ) Recuperado 5 de julio de 2010,
de http://www.talkorigins.org/faqs/evolphil/naturalism.html
[lxi]
Gilean A. T. McVean and Brian Charlesworth, The Effects of Hill-Robertson Interference Between Weakly Selected
Mutations on Patterns of Molecular Evolution and Variation, Genetics Society of America, Genetics 155: 929–944 (2000)
Recuperado 5 de julio de 2010, de http://www.genetics.org/cgi/content/full/155/2/929
[lxii]
Joachim Moll, Patrizia Barzaghi, Shuo Lin, Gabriela Bezakova, Hanns Lochmu ller, Eva Engvall, Ulrich Muller & Markus A.
Ruegg, An agrinminigene rescues dystrophic symptoms in a mousemodel for congenital muscular dystrophy, Nature 413, ( 2001)
Recuperado 5 de julio de 2010, de http://www.nature.com/nature/journal/v413/n6853/abs/413302a0.html
[lxxiii]
Wong, J. T.-F. A co-evolution theory of the genetic code,Proc. Natl. Acad. Sci. USA 72, 1909-1912 (1975)
[lxxviii]
J. G. Blank, M. G. Knize, and Glen Nakafuji, Laboratory Studies of Shock-Induced Amino Acid Polymerization and Implications for Extraterrestrial Delivery and Impact-Generated Production of Biomolecules, Lawrence Berkeley National Laboratory, Recuperado 5 de julio de 2010, de http://www.lpi.usra.edu/meetings/lpsc2002/pdf/2075.pdf
[lxxxi]
Elizabeth T. N. Bui, Peter J. Bradley, And Patricia J. Johnson, A common evolutionary origin for mitochondria and hydrogenosomes, Proc. Natl. Acad. Sci. USA Vol. 93, pp. 9651–9656 (1996) Recuperado 5 de julio de 2010, de http://www.pnas.org/content/93/18/9651.full.pdf
_________________________________________________
Título Original de la obra:
Biología de sistemas: una mirada a lo más profundo de nuestro origen, evolución y funcionamiento. Copyright © 2003-2010 por la ANUIES/SINED/UMSNH. CIE Tzintzuntzán No. 173, Col. Matamoros C.P. 58240, Edificio E Planta alta, Morelia Michoacán, México. Tel/fax: 3-14-28-09
Ésta fue publicada originalmente en Internet bajo la categoría de contenido abierto sobre la URL: http://dieumsnh.qfb.umich.mx/ mismo título y versión de contenido digital. Este es un trabajo de autoría publicado sobre Internet Copyright © 2003-2010 por la Coordinación de Innovación Educativa Q.F.B./UMSNH, protegido por las leyes de derechos de propiedad de los Estados Unidos Mexicanos. No puede ser reproducido, copiado, publicado, prestado a otras personas o entidades sin el permiso explícito por escrito de la ANUIES/SINED/UMSNH o por los Autores.
Ochoa, H. E., et al (2010) Biología de sistemas. Una mirada a lo más profundo de nuestro origen, evolución y funcionamiento. México, ANUIES/SINED/UMSNH. vii, 302 p.; carta
Publicado en URL: http://dieumsnh.qfb.umich.mx/
eohqfb@yahoo.com.mx
Así, estimado lector, asegúrese de contestar para cada elemento el "¿cómo?", pregunta que infiere: ¿qué significa el texto?, ¿qué significa cada pieza?, esto le ayudará a mostrar cómo se entiende el texto desde su analítica de piezas, lo que derivará en un reporte de lectura, desde donde se analiza con preponderancia la evidencia para cada elemento de la prosa. En este texto, debido a su extensión no atendemos los diferentes protocolos en los que se reporta la lectura, pero están a su disposición en la URL: http://www.libertadacademica.com/conalepweb2013/ObjetoReportesLectura/index.html.
Coordinación de innovación educativa LAN UMSNH
- Página principal CIE
- Texto academico
- Diplomado tutor escritor
- Cursos en línea




{kind=link}